创新背景
人类所熟悉的动植物的有性生殖过程需要卵细胞和精子细胞结合,精子和卵子的融合标志着受精的高潮,也标志着胚胎发育的开始。
创新过程
以往的科学家研究证明有性生殖的出现可以追溯到大约10亿至20亿年前,以色列理工学院的研究人员与乌拉圭、瑞士、瑞典、法国、英国和阿根廷的研究人员合作推测,允许这种聚变事件发生的机制早在30亿年前就出现了。相关研究成果论文《发现与真核HAP2/GCS1配子融合蛋白同源的古菌融合素》发表在2022年7月6日出版的《自然通讯》杂志上。
由于两个细胞都包含了后代所需的恰好一半的遗传信息,多个精子与一个卵子的异常融合将会导致异常的发育。因此,这一过程受到严格监管。被称为“fusogens”的特殊蛋白质必须在精确的时间和地点出现,使卵子和精子融合成一个。实验室研究了几种生物中的fusogens,并首先在线虫秀丽隐杆线虫中鉴定表征了两种这样的蛋白质。这些蛋白质参与器官发育,但不参与受精。令人惊讶的是,结构分析显示这些蛋白与植物中另一个参与受精的融合素(GCS1/HAP2)具有非常相似的三维结构。这种结构相似的融合蛋白家族被命名为Fusexin,在植物、动物、病毒、单细胞藻类和寄生虫中有代表性。

为了扩展和描述Fusexin家族的起源和进化,研究人员对来自不同环境的基因序列进行了生物信息学研究。在从土壤、盐湖、淡水和海洋沉积物中筛选样本后,他们发现了属于古菌的96个序列,这些序列与已知的融合蛋白有一些相似之处。这些序列被命名为fusexin1 (Fsx1),专家团队证实,它们属于古细菌物种,据估计起源于30亿年前。然而,目前尚不清楚Fsx1编码的蛋白质是否看起来与fusexin家族成员相似,以及它是否真正能够介导细胞间融合。
为了确定Fsx1蛋白的结构,瑞典卡罗林斯卡研究所的研究人员使用晶体学方法破译了Fsx1蛋白的三维构象。Fsx1蛋白包含三个结构域,与已知的Fusexin成员非常相似,并与其他已知的fusogens一样排列在一个三段复合体(称为三聚体)中。并且,Fsx1还有一个在任何已知的Fusexin成员中都没有的额外的第四个域。
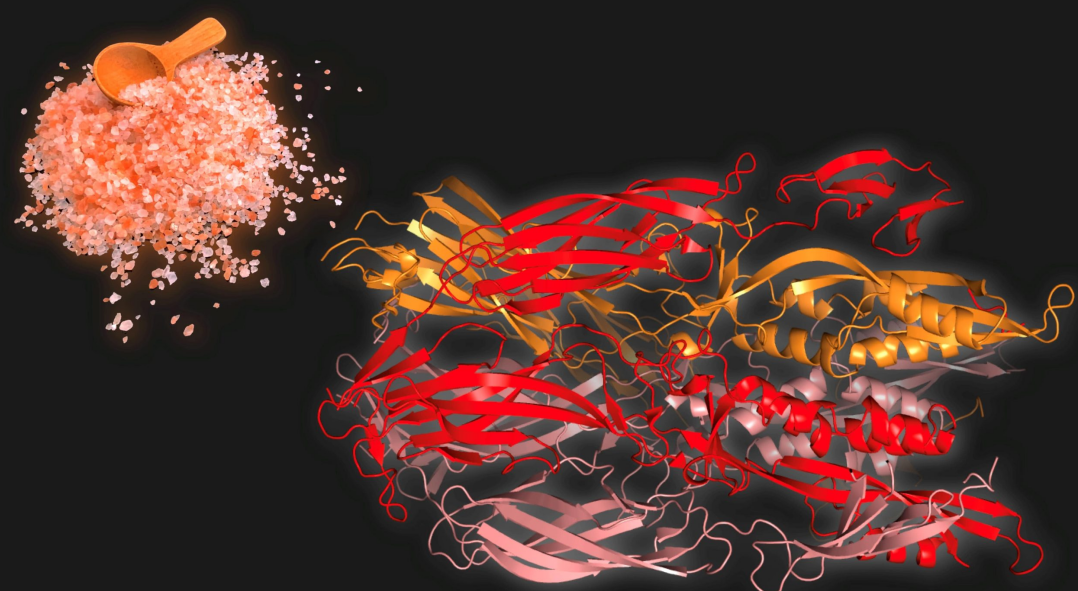
为了证明Fsx1蛋白具有融合蛋白的作用,研究人员在来自哺乳动物的细胞培养中表达了Fsx1蛋白,这些细胞培养物通常不会融合。实验表明来自古菌的Fsx1确实诱导了这些在10亿至20亿年前分化的哺乳动物细胞的融合。
已知病毒中的融合蛋白介导病毒进入宿主细胞,而在真核生物中,它们在器官雕刻、神经元修复和性方面发挥作用。但是融合蛋白和病毒谁先进入细胞尚无定论,研究人员在一个融合体被病毒捕获用于有性繁殖和一个病毒蛋白被植物采用的情况外提出了第三种情况:所有的融合蛋白都起源于古生菌,从那里谱系分裂成各种功能,从病毒感染到精子和卵子融合,比有性生殖早了10亿年。
下一步重要的工作是研究Fsx1蛋白在自然界中的作用,探究它们是否会像植物和动物的融合蛋白一样融合古生细胞,融合配子(例如卵子和精子)来促进性别相似的DNA交换。此外,研究还需要进行平行研究来了解Fsx1蛋白和GCS1/HAP2蛋白之间的进化历史,以确定它们的起源。
古菌融合素和其他尚未被发现的融合素可能有助于理解细胞是如何从表面上简单的形式,共享各自的DNA片段,进化到今天的复杂生命形式,经历有性繁殖。因此,古生菌等古老生物也可能含有融合蛋白的发现现在提出了一种有趣的可能性,即Fsx1蛋白是病毒、植物和动物融合蛋白的祖先版本。
创新关键点
研究通过对来自不同环境的基因序列进行生物信息研究,证明Fsx1蛋白具有融合蛋白的作用,合理推测精子细胞和卵细胞融合的细胞机制起源于30亿年前。
智能推荐
PABP/R-motif介导的蛋白重编程新机制
2022-08-04该研究揭示了不同于经典的eIF2α磷酸化介导的蛋白重编程机制,同时因为R-motif同样广泛存在于果蝇、小鼠和人体的胁迫相关mRNA的5’端前导序列中,本研究发现的蛋白翻译起始调控机制可能同样广泛存在于其他真核生物体中。
涉及学科涉及领域研究方向STAND atp酶蛋白通过识别病毒三维结构对抗病毒感染
2022-08-14研究团队在本研究中整合了遗传学、生物信息学、生物化学和结构生物学的方法,发现了原核生物中的STAND atp酶蛋白能够直接感知病毒中的门户蛋白和末端酶,并通过切断细菌自身的DNA,杀死细胞,达到防御感染的效果。
涉及学科涉及领域研究方向生命科学理论创新 | 研究人员揭示延长植物蛋白寿命的细胞机制
2022-09-02通过使用拟南芥在蛋白质组中寻找HYPK,测试控制蛋白质寿命机制的关键蛋白质。
涉及学科涉及领域研究方向创新利用TadA8e进入双链DNA进行碱基转换
2022-06-29跳出思维局限,创新利用TadA8e进入双链DNA进行碱基转换。
涉及学科涉及领域研究方向